|
Postmolt (postecdysis)
Postmolt is the stage just following exuviation (shedding of the old exoskeleton).
It is the period when the exoskeleton expands due to increased hemolymph volume from water influx. Water influx occurs across the epidermis, gills, and gut. After several hours or days (depending upon total length of the molt cycle), the new exoskeleton hardens and retains its rigidity (Chang, 1992).
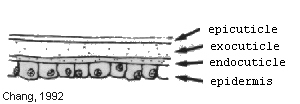 Immediately after ecdysis, the only layers present are the epicuticle and exocuticle. Within a few hours the epidermis starts to secrete the endocuticle. Most of the cuticle must be derived from materials stored in the epidermis, as feeding does not begin until the prawn is well into the intermolt stage. This secretion continues until the prawns are in the intermoult condition, when the three layers are fully formed (Dall et al., 1990).
Immediately after ecdysis, the only layers present are the epicuticle and exocuticle. Within a few hours the epidermis starts to secrete the endocuticle. Most of the cuticle must be derived from materials stored in the epidermis, as feeding does not begin until the prawn is well into the intermolt stage. This secretion continues until the prawns are in the intermoult condition, when the three layers are fully formed (Dall et al., 1990).
Intermoult
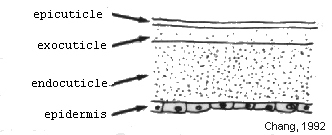 During intermolt the exoskeleton becomes much harder through mineral and protein deposition. Shrimp exoskeleton is relatively thin and soft compared to crabs and lobsters (Chang, 1992). The volume as well as the weight of the whole prawn increases by 3-4% during the intermoult period. This increase may be due to extension of the thin intersegmental connections of the abdomen, and supports the concept that growth in penaeids is a more continuous process than in the heavily armoured decapods, which moult relatively infrequently (Dall et al., 1990). During intermolt the exoskeleton becomes much harder through mineral and protein deposition. Shrimp exoskeleton is relatively thin and soft compared to crabs and lobsters (Chang, 1992). The volume as well as the weight of the whole prawn increases by 3-4% during the intermoult period. This increase may be due to extension of the thin intersegmental connections of the abdomen, and supports the concept that growth in penaeids is a more continuous process than in the heavily armoured decapods, which moult relatively infrequently (Dall et al., 1990).
Premolt (proecdysis)
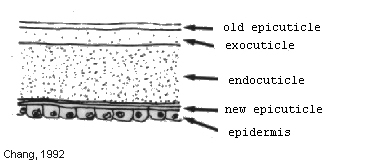 Premolt (proecdysis) stage occurs just prior to exuviation, and is characterized by separation of the old exoskeleton from the underlying epidermal layer. The old exoskeleton is partly reabsorbed, and energy reserves are mobilized from the midgut gland. Premolt begins with an increase in concentration of molting hormone in the hemolymph (Chang, 1992). Premolt (proecdysis) stage occurs just prior to exuviation, and is characterized by separation of the old exoskeleton from the underlying epidermal layer. The old exoskeleton is partly reabsorbed, and energy reserves are mobilized from the midgut gland. Premolt begins with an increase in concentration of molting hormone in the hemolymph (Chang, 1992).
The first indication that the prawn is entering proecdysis is the withdrawal of the epidermis from the old cuticle (apolysis). Later the epidermis starts to hypertrophy and cells, which appear to have a storage function, accumulate in it. As the prawn proceeds through this stage, the epidermis starts to secrete a new epicuticle and exocuticle.
Feeding starts to decline and has completely ceased by the end of the proecdysis. Thus reserves need to be available for cuticle synthesis and for the period when feeding ceases. The material for cuticle synthesis are derived from two sources: accumulated reserves due to feeding and resorption from the old cuticle (Dall et al., 1990).
Ecdysis
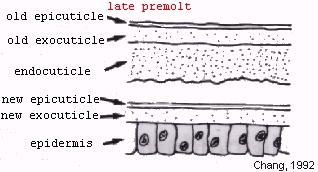 Ecdysis, as a stage, only lasts a few minutes. It begins with the old exoskeleton opening at the dorsal junction of the thorax and abdomen in decapod crustaceans, and is completed when the animal escapes from its confines (Chang, 1992).
Ecdysis, as a stage, only lasts a few minutes. It begins with the old exoskeleton opening at the dorsal junction of the thorax and abdomen in decapod crustaceans, and is completed when the animal escapes from its confines (Chang, 1992).
Since uptake of water determines the size until the next moult, this is the most important single physiological process at the end of the proecdysis and during ecdysis. Other decapods absorb most of their water by drinking, but this process has not been investigated for Penaeidae. Presumably there is some uptake in the late proecdysis, but it appears that water uptake must be limited to a few housrs on either side of ecdysis, as the cephalothorax swells only immediately before ecdysis, and the cuticle hardens appreciably within a few hours afterwards (Dall et al., 1990).
|
